关于韧皮部运输机理的研究通常包括以下几个方面的内容:
(1)韧皮部运输的速度和方向;(2)韧皮部运输的动力;(3)同化物从叶肉细胞进入筛管的机理和调节;(4)同化物在筛管中运输的机理;(5)同化物从筛管向库细胞的释放机理;(6)影响上述这些过程的因素等。
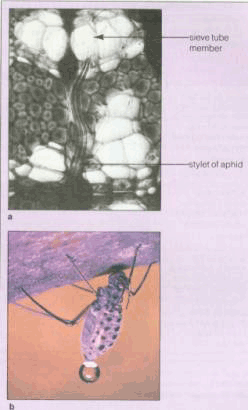
| 一、研究韧皮部同化物运输的方法 要研究韧皮部运输的机理,首先需了解韧皮部运输的基本情况、同化物的种类、浓度及单位时间单位运输面积通过同化物的量等。常采用的研究方法有以下几种: (一)收集韧皮部汁液的方法 收集韧皮部汁液的方法有多种,其中一个比较成功的方法是利用刺吸式昆虫的口器吻针法。例如蚜虫的口
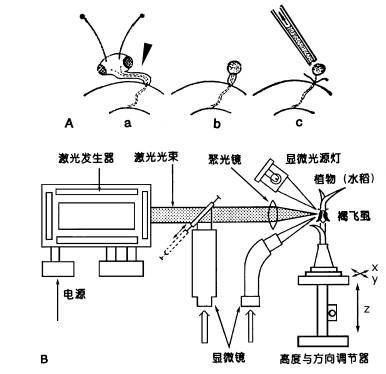
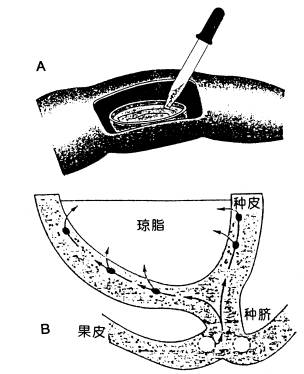
A.用蚜虫口器收集筛管汁液的示意图。 a.将蚜虫的吻针连同下唇一起切下; b.切口溢出筛管汁液; c.用毛细管收集溢泌液。B.用激光切断飞虱口针的装置。用显微镜观察与聚焦,当焦点聚在飞虱口针时,开启激光器,随即口针被烧断。 器可以分泌果胶酶帮助其吻针刺入韧皮部筛管分子,当蚜虫的吻针刺入筛管分子后,用CO2将其麻醉,切除母体而留下吻针。由于筛管正压力的存在,韧皮部汁液可以持续不断地从吻针流出(图6-8A)。有报道,在柳树上用此法可以连续收集韧皮部汁液(4μl·h-1)达4d之久。在此基础上人们已设计出了用激光切断飞虱口针(图6-8B),用压力探针来收集水稻韧皮部汁液的方法。 另一种收集韧皮部汁液的方法是在韧皮部上切一个1mm深的刀口,然后用毛细管收集韧皮部汁液。该法仅适用于韧皮部和木质部相对独立的植物,如木本植物、棉花、麻类等。许多植物在切口处常出现胼胝质的堵塞现象。若用金属离子螯合剂EDTA或EGTA溶液处理切口,可以避免这种堵塞现象,这是因为胼胝质合成酶的活化需要Ca2 ,而EDTA能与Ca2 螯合, 阻止胼胝质的形成。
A.用解剖刀将部分豆荚壳切除,开一“窗口”,切除正在生长种子的一半(远种脐端),将另一半种子内的胚性组织去除,仅留下种皮组织和母体相连部分,制成空种皮杯。在空种皮杯中放入4%琼脂或含有EGTA溶液的棉球,收集空种皮中的分泌物; B.同化物在空种皮杯中卸出的途径。(Hopkins,1995) (二)测定韧皮部运输速度的方法 目前应用比较普遍的方法有两种。一种是利用染料分子作为示踪物,用显微注射技术将染料分子直接注入筛管分子内,追踪染料分子在筛管中的运输状况,根据单位时间中染料分子的移动距离来计算运输速度。另一种是放射性同位素示踪技术,常用的同位素是14C。将14CO2气体饲喂叶片,然后追踪14C标记的碳水化合物在筛管中的运输状况、运输速度,用这种技术还可研究同化物的分配动态。可以说有关韧皮部运输的许多信息均来自于同位素示踪技术的应用。 (三)新技术的应用 近年来,新技术包括分子生物学技术被广泛地应用到韧皮部运输的研究中,其中已被应用的技术有:(1)用共聚焦激光扫描显微镜(confocal laser scanning microscope,CLSM)直接观察完整植株体内韧皮部同化物运输(包括韧皮部装卸)的影像。(2)空种皮技术(empty seed coat technique,emptyovule technique),此法适用于豆科植物(图6-9)。实验证明,在短时间内,空种皮杯内韧皮部汁液的收集量与种子实际生长量相仿, 借此用以研究同化物韧皮部卸出机理和调节;(3)微注射法(microinjection technique),用微量进样器将少量激素等化学物质注入到正在生长的种子中,观察与测定激素等化学物质对种皮卸出同化物的影响;(4)应用分子生物学技术将编码绿色荧光蛋白(green fluorescent protein, GFP)的基因导入病毒基因组内,这样可直接观察病毒蛋白在韧皮部中的运输。 |
| 二、韧皮部中运输的物质 韧皮部汁液化学组成和含量因植物的种类、发育阶段、生理生态环境等因素的变化而表现出很大的变异。一般来说,典型的韧皮部汁液样品其干物质含量占10%~25%,其中多数是糖,其余为蛋白质、氨基酸、无机和有机离子,且汁液中的氨基酸主要是谷氨酸和天冬氨酸。有些植物韧皮部汁液样品中还含有植物内源激素,如生长素、赤霉素、细胞分裂素和脱落酸。
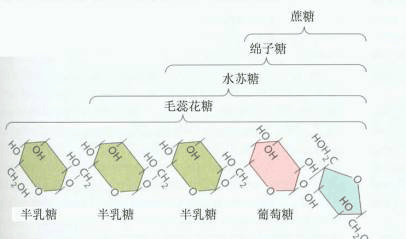
在多数植物中蔗糖是韧皮部运输物的主要形式。少数科的植物韧皮部汁液中除蔗糖外,还含有棉子糖、水苏糖、毛蕊花糖等(图6-11), 有的还含有糖醇,如甘露醇、山梨醇等。
图 6-11 韧皮部汁液中几种糖的结构 为什么蔗糖是韧皮部运输物质的主要形式?其一可能是蔗糖及其它一些寡聚糖是非还原糖,它们在化学性质上具有较还原糖更大的稳定性;其二可能是蔗糖水解时能产生相对高的自由能。由于蔗糖是一种分子小、移动性大,化学性质稳定,且含有高水解自由能的化合物,因此它适合进行长距离的韧皮部运输。 |
| 三、同化物运输的方向和速度 同化物运输(assimilate transportation)的方向决定于源和库的相对位置。一般来说,韧皮部内同化物运输的方向是从源器官向库器官运输。然而,在植物生长发育的不同阶段,常同时存在多个源和多个库,所以由一个源器官制造的同化物常需同时供应多个库器官所需,一个库器官也可能接纳多个源器官供应的同化物,而且这些源库常分布于植株的不同位置。因此,同化物既可能向顶也可能向基运输,这种韧皮部同化物的双向运输(bidirectional transport)已被许多实验证实。然而对某一个筛管来说,通常认为同化物在其中的运输是单向的,而不是双向的。同化物运输的速度一般为0.2~2m·h-1。不同植物或不同生长势的植物个体,其同化物的运输速度不一样,生长势大的个体运输速度快。同化物运输速度是韧皮部物质运输的一个重要指标,然而,人们往往对其中运输的物质的量更感兴趣。因此,提出了比集转运速率(specific mass transfer rate, SMTR)的概念,即单位时间单位韧皮部或筛管横切面积上所运转的干物质的量。 SMTR(g·cm-2·h-1)= 运转的干物质量 韧皮部(筛管)横切面积×时间 (6-1) 或 SMTR(g·cm-2·h-1 )=运输速度×运转物浓度 (6-2) 其中运输速度以cm·h-1、运转物浓度以g·cm-3表示。如马铃薯块茎与植株地上部由韧皮部横切面为0.004cm2的地下蔓相连,块茎在50d内增重230g,块茎含水量为75%,则此株马铃薯同化物运输的比集转运速率为: SMTR= 230×(1-75%) 0.004×24×50 ≈12(g·cm-2·h-1) 多数植物韧皮部的SMTR为1~13 g·cm-2·h-1 ,最高可达200 g·cm-1·h-1。筛管分子横切面一般占整个韧皮部横切面的20%,上述韧皮部中筛管的SMTR则为60 g·cm-2·h-1。
图 6-12 源叶中韧皮部装载途径示意图 |
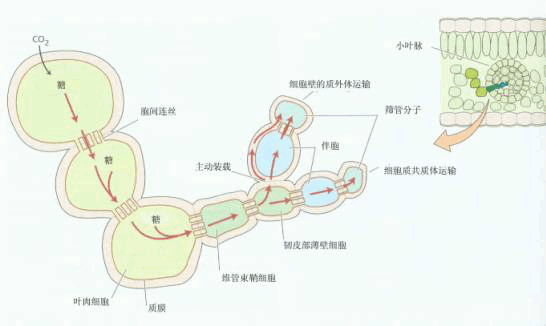
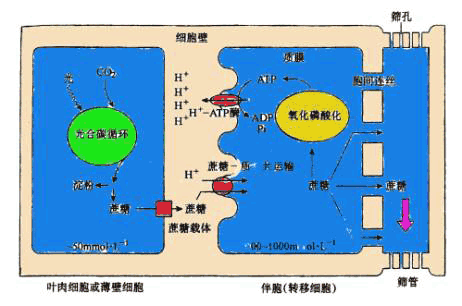
| 四、韧皮部装载 韧皮部装载(phloem loading)是指同化物从合成部位通过共质体或质外体的胞间运输进入筛管的过程。而同化物从韧皮部薄壁细胞进入伴胞和筛管的过程则特指为筛管装载(sieve loading)。韧皮部装载途径如图6-12所示。 整个途径由三个区域组成,即光合同化物生产区、累积区和输出区。同化物生产区为叶肉细胞(C4植物还会含维管束鞘细胞)。叶绿体进行光合作用形成的磷酸丙糖通过叶绿体膜上的磷酸转运器(Pitranslocator, PT)进入细胞质,在一系列酶作用下合成蔗糖,并通过胞间运输进入累积区。累积区主要由小叶脉末端的韧皮部的薄壁细胞组成,而输出区则主要是指叶脉中的SE-CC。从对整个叶片而言,同化物累积区和输出区是一个难以分割的连续体系。 (一)装载途径 同化物从周围的叶肉细胞转运进韧皮部SE-CC复合体的过程中存在着两种装载途径——质外体途径和共质体途径。 1.质外体装载 质外体装载 (apoplasmic phloem loading)是指光合细胞输出的蔗糖进入质外体,然后通过位于SE-CC复合体质膜上的蔗糖载体逆浓度梯度进入伴胞,最后进入筛管的过程。跨膜质子梯度是由ATPase水解ATP移动H 所形成的(图6-13)。 支持质外体装载的实验证据主要来自以下几方面: ①许多植物(如大豆,玉米)小叶脉SE-CC复合体与周围薄壁细胞间无胞间连丝连接 ②在SE-CC复合体介面上存在大的渗透梯度,SE-CC内的蔗糖浓度可高达800~1 000mmol·L-1,而叶肉细胞的蔗糖浓度只有50mmol·L-1左右 ③用14C标记的大豆叶片,质外体中存在高浓度的14C蔗糖。质外体中蔗糖含量占细胞总蔗糖含量的7% ④用14C蔗糖和14C葡萄糖进行的放射性自显影研究表明,SE-CC复合体可以直接吸收蔗糖,但不吸收葡萄糖等非运输形式的糖分子 ⑤代谢抑制剂如DNP及厌氧处理会抑制SE-CC复合体对蔗糖的吸收,这表明质外体装载是一个主动过程 ⑥用质外体运输抑制剂对氯汞苯磺酸(parachloro-mercuribenzene sulfonate)处理14CO2标记的叶片,然后进行放射性自显影,发现SE-CC复合体中几乎无14C蔗糖存在。这些结果都直接或间接地说明韧皮部装载可通过质外体途径进行。 2.共质体装载 共质体装载(symplastic phloem loading)途径是指光合细胞输出的蔗糖通过胞间连丝顺蔗糖浓度梯度进入伴胞或中间细胞,最后进入筛管的过程(图6-12)。支持共质体装载途径的实验证据有:①许多植物叶片SE-CC复合体和周围薄壁细胞间存在紧密的胞间连丝连接 ②一些植物同化物的韧皮部装载对PCMBS不敏感 ③将不能透过膜的染料如荧光黄CH(Lucifer yellow CH)注入叶肉细胞,一段时间后可检测到筛管分子中存在这些染料。 另外,一些植物同化物的韧皮部装载既可以通过质外体途径,也可以通过共质体途径。如西葫芦, 其叶片支脉的伴胞既有普通型的,也有属于中间细胞的,推测前者装载蔗糖是通过质外体途径,而后者则是通过共质体途径。总之,这两种途径的相对比例因植物种类而异,且随叶片的发育阶段、昼夜和季节的变化以及生态条件的变化而变化。 (二)装载机理 由于韧皮部装载有多种模式,因此相应的装载机理也不同。 1.质外体装载机理 质外体装载过程包括以下三个阶段。 (1)同化物从光合细胞向质外体的释放 参与释放过程的细胞内同化物库的大小是决定同化物从光合细胞向质外体释放能力的一个主要因素。并不是所有光合细胞都参与同化物的释放过程,而只有那些临近SE-CC复合体的光合细胞才能直接参与该过程。因此,大部分光合细胞形成的光合同化物首先经胞间连丝进入这些特化细胞,然后再通过这些细胞释放到质外体中。 (2)同化物在质外体的去向 20世纪80年代以前,一般认为蔗糖进入质外体后在蔗糖酶作用下先水解形成葡萄糖和果糖,然后再进入SE-CC复合体。然而,以后许多实验结果表明,蔗糖进入质外体后并不发生进一步的代谢或修饰,而是直接以蔗糖分子进入SE-CC复合体。用14C蔗糖和葡萄糖喂甜菜叶片,其结果是在筛管中只检测到蔗糖的存在。
图 6-13 蔗糖从叶肉细胞向筛管的质外体装载 在叶肉细胞质合成的蔗糖,经质膜上的载体进入质外体,运至伴胞(转移细胞),质膜外的质子蔗糖共运输蛋白在H 梯度的驱动下,进入伴胞(或转移细胞),然后经胞间连丝进入筛管。伴胞(或转移细胞)质膜外H 梯度的建立,依赖ATPase分解ATP的反应,而 ATP来自蔗糖分解后的氧化磷酸化。 (前岛正义,1995) (3) 同化物进入SE-CC复合体 蔗糖装载是通过SE-CC复合体的伴胞进行的,蔗糖进入伴胞是通过位于质膜上的蔗糖质子共运输蛋白(sucroseH symporter),它运输蔗糖时与质子传递相偶联。ATP在质膜ATPase作用下产生跨膜的质子梯度,由跨膜的质子梯度驱动蔗糖的跨膜运输(图6-13)。蔗糖和质子共运输的化学准量关系(stoichemistry)随蔗糖浓度的变化而变化,一般在蔗糖浓度低于5mmol·L-1 时,每吸收2mol蔗糖偶联吸收1mol H (即2∶1),但当蔗糖浓度在5~15 mmol·L-1 之间时,蔗糖和质子偶联共运输的化学准量关系为6∶1。蔗糖是中性分子,蔗糖和质子的偶联共运输将引起膜势的去极化(depolarization), 但去极化作用只是瞬间的,因为在ATPase作用下质子将以更高的速度从伴胞泵出,从而补偿了由于质子与蔗糖的共运输而引起的质外体内质子浓度的降低。一些证据支持质子与蔗糖的共运输的假说,如韧皮部汁液的pH比质外体中高,韧皮部汁液中含有ATP,以及SE-CC复合体的质膜常保持负的膜电势。 2.共质体装载机理 在进行共质体装载的植物中,筛管中糖的主要运输形式是寡聚糖(棉子糖、水苏糖、毛蕊花糖等)和蔗糖。早期对同化物共质体装载持反对观点的主要理由是这种装载模式违反了热力学规律,即SE-CC复合体逆浓度梯度积累糖类。目前虽然对共质体装载机理仍争论不休,但有两种假说可部分解释这种逆浓度梯度积累糖类的机理。第一种假说称为多聚化截留机理(polymerization trap mechanism)。该假说认为,叶肉细胞内形成的蔗糖和肌醇半露糖苷顺着其浓度梯度扩散进入中间细胞,在那里合成棉子糖或水苏糖或毛蕊花糖,由于这些寡聚糖分子的直径超过了连接叶肉细胞和中间细胞间的胞间连丝横断面的直径,从而阻止了糖向叶肉细胞的倒流(reverseflow),而中间细胞与筛分子间的有分支胞间连丝的直径足以让这些糖分子通过而进入筛管。支持该假说的实验证据有:以南瓜为材料,在其叶片中检测到棉子糖-6-半露糖苷转移酶(也称水苏糖合成酶),在中间细胞中检测到寡聚糖的存在,在叶脉中检测到大量寡聚糖的存在。第二种假说称为内质网络运输机理(endoplasmic network transport mechanism),该假说认为,寡聚糖主要是在叶肉细胞内形成的,并在那儿被装载到横穿连接叶肉细胞和中间细胞胞间连丝的液泡-维管状网络结构内,然后通过扩散作用进入筛管分子。 |
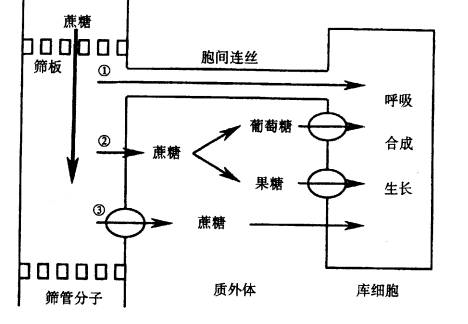
| 五、韧皮部卸出 光合同化物一旦装载进入筛管,就会沿整个韧皮部运输途径不断地和周围细胞进行物质交换,即卸出和再装载,其中韧皮部的卸出主要发生在库端。韧皮部卸出(phloem unloading)是指光合同化物从SE-CC复合体进入库细胞的过程。韧皮部卸出对同化物的运输、分配以及作物最终经济产量等都起着极其重要的调节作用。 (一)卸出途径 韧皮部卸出的途径有两条:一条是共质体途径;另一条是质外体途径。
图6-14 由韧皮部向库细胞卸出同化物的途径 ①共质体卸出; ②③质外体卸出 1. 共质体卸出 一般来说,正在生长发育的叶片和根系,同化物是经共质体途径卸出的,即蔗糖通过胞间连丝沿蔗糖浓度梯度从SE-CC复合体释放到库细胞的代谢部位(图6-14①)。 支持这一观点的实验证据主要有: (1)膜不通透染料的卸出研究 用膜不通透染料本carboxyfluorescein (CF)观察拟南芥根尖韧皮部卸出途径,证明CF是通过胞间连丝运输的。 (2)14C同位素示踪研究 同化物经共质体途径卸出最直接的证据是运用14C蔗糖进行的研究。在14C蔗糖从韧皮部卸出的过程中,没有检测到14C蔗糖进入质外体空间或在胞外降解,这也有力地支持了共质体卸出这种途径。 (3)溶质跨膜运输抑制剂的应用 运用蔗糖跨膜运输抑制剂PCMBS发现,PCMBS并不抑制蔗糖的韧皮部卸出,这说明蔗糖的韧皮部卸出是通过共质体途径进行。 2.质外体卸出 某些植物或组织(如甜菜的块根、甘蔗的茎及种子和果实等),其韧皮部卸出是通过质外体途径进行的。这些植物组织的SE-CC复合体与库细胞间通常不存在胞间连丝,所以SE-CC复合体中的蔗糖只能通过扩散作用或通过膜上的载体先进入质外体空间,然后直接进入库细胞(图6-14③),或降解成单糖后再进入库细胞(图6-14②)。 根据蔗糖进入质外体后的命运,又可将同化物韧皮部卸出的质外体途径分成两条子途径: 子途径一是SE-CC复合体中的蔗糖通过扩散作用跨细胞膜进入质外体空间。因为这一过程对代谢抑制剂和PCMBS不敏感,因此,不涉及需要能量活化的载体的参与。但是,保持扩散作用不断进行需要膜两侧维持足够高的蔗糖浓度梯度,这不仅要求光合细胞具有高的光合速率,而且要求进入质外体空间内的蔗糖不断地被移走(即或被降解或被其他细胞吸收)。许多证据表明,细胞壁转化酶(invertase)是催化质外体空间内蔗糖降解的主要酶,而水解形成的葡萄糖和果糖就可被库细胞主动吸收。在玉米、高粱、粟等植物的种子内,同化物的韧皮部卸出是通过这条途径进行的。 子途径二是SE-CC复合体内的蔗糖释放到质外体空间并由位于膜上的载体介导。该途径会受低温、代谢抑制剂和PCMBS的抑制,这表明蔗糖通过这条途径卸出需要有能量供应。这条途径在豆科类植物种子中普遍存在。 (二)韧皮部卸出的机理及调节 1.共质体卸出 同化物通过共质体途径卸出存在两种情况:一种是通过扩散作用(diffusion);另一种是通过集流方式。通过扩散作用卸出的速率取决于胞间连丝两侧被运输同化物的浓度梯度和胞间连丝的传导能力。根据菲克定律,同化物通过共质体途径卸出的速率(Rd): Rd=n〔DA(C -Cs)/l〕 (6-3) 上式中n-单位界面上胞间连丝的数目;D-同化物在胞间连丝通道中的扩散系数;A-胞间连丝横切面积;C 和Cs-筛管和库细胞中同化物的浓度;l-胞间连丝通道的有效长度。 由(6-3)式可知,对某一特定的器官(即n、 A、 l为一定值),同化物共质体卸出的速率与胞间连丝的传导能力和胞间连丝两侧被运输同化物的浓度梯度成正比。许多证据表明这种共质体卸出方式的存在,如在玉米根尖和小麦籽粒内检测到筛管与库细胞间存在大的蔗糖浓度梯度。 若共质体卸出通过集流方式进行,则同化物卸出速度除取决于胞间连丝的传导能力外,还与胞间连丝两侧的压力梯度有关。这种卸出方式可用试验来证明。如将库细胞放入含不能透过膜的渗透介质溶液中,卸出速率显著增加,并且与所用溶液渗透势的大小呈线性相关。对胞间连丝显微结构的观察以及胞间连丝两侧压力梯度的测定也证明许多植物或器官韧皮部卸出是通过集流方式进行的。 有关韧皮部卸出调节机理的研究主要集中在胞间连丝的结构和胞间连丝两侧压力或浓度梯度方面。单位输导面积上胞间连丝的数目以及胞间连丝的超微结构受发育调节,因而对韧皮部卸出的调节是长期效应。然而,胞间连丝的传导能力、同化物浓度和细胞膨压的变化对韧皮部卸出具有即时的调节作用。许多实验证明,库细胞接纳和转化同化物的能力直接决定了胞间连丝两侧同化物浓度或压力差,库细胞中催化蔗糖降解的酶(转化酶和蔗糖合成酶)活性与同化物进入库细胞的速度存在密切的关系。 胞间连丝的传导能力是控制卸出速度的另一个主要因素。胞间连丝的超微结构和传导能力不是固定不变的,而是受到生理条件、发育阶段和环境条件的影响,此外,低温、水分胁迫都会降低胞间连丝的传导能力。 2.质外体卸出 蔗糖从SE-CC复合体跨质膜进入质外体是通过顺浓度梯度的简单扩散作用进行的,而蔗糖跨质膜从质外体进入库细胞是由载体介导并需要消耗能量的质子蔗糖共运输机制进行的。因此,蔗糖从SE-CC复合体内通过质外体途径卸出速度决定于蔗糖质子共运输的速度。而决定蔗糖质子共运输速度的共运输蛋白的活性受膨压调节,虽然其确切的调节机理有待进一步研究,但有结果显示,它是通过调节H ATPase活性,从而影响与蔗糖吸收密切相关的质子动力势而实现的。 |
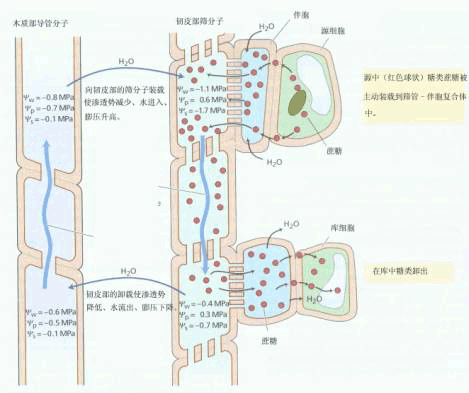
| 六、韧皮部同化物运输的机理 1930年明希(E.Münch)提出了解释韧皮部同化物运输的压力流学说(pressure flow hypo thesis)。该学说的基本论点是,同化物在筛管内是随液流流动的,而液流的流动是由输导系统两端的膨压差引起的。自该学说提出以来,许多学者都致力于能更完整、更正确地解释同化物韧皮部运输的现象,也曾提出过多种假说,如简单扩散作用、细胞质环流(cyclosis)、电渗流动、收缩蛋白、离子泵等假说,然而这些假说后来均被实验证明是不完整的或错误的。
图6-15 韧皮部中同化物运输分配的压力流动模式图 木质部和韧皮部中可能的Ψw、Ψp、Ψs数值都在图中说明。 目前被人们广为接受的学说是在明希最初提出的压力流学说基础上经过补充的新的压力流学说。新学说认为,同化物在筛管内运输是一种集流,它是由源库两侧SE-CC复合体内渗透作用所形成的压力梯度所驱动的(图6-15)。而压力梯度的形成则是由于源端光合同化物不断向SE-CC复合体进行装载,库端同化物不断从SE-CC复合体卸出,以及韧皮部和木质部之间水分的不断再循环所致。即光合细胞制造的光合产物在能量的驱动下主动装载进入筛管分子,从而降低了源端筛管内的水势,而筛管分子又从邻近的木质部吸收水分,以引起筛管膨压的增加;与此同时,库端筛管中的同化物不断卸出并进入周围的库细胞,这样就使筛管内水势提高,水分可流向邻近的木质部,从而引起库端筛管内膨压的降低。因此,只要源端光合同化物的韧皮部装载和库端光合同化物的卸出过程不断进行,源库间就能维持一定的压力梯度,在此梯度下,光合同化物可源源不断地由源端向库端运输。 根据压力流模型可预测韧皮部的运输应具有如下特点:① .各种溶质以相似的速度被运输;②.在一个筛管中运输是单方向的;③.筛板的筛孔是畅通的;④.在筛管的源端与库端间必须有足够大的压力梯度;⑤.装载与卸出过程需要能量,而在运输途中不需消耗大量的能量。 关于前三点,曾有许多实验结果与其不符。 测定不同示踪物如14C蔗糖、32P磷酸根和3H2O在韧皮部中的运输速度,发现蔗糖速度最快,磷酸根次之,H2O最慢,这似乎与压力流学说是不相符的。但有以下可能性:①水很容易透过膜,它可以由筛管进入周围组织,周围组织中的水又进入筛管,水在运输途中走了一条“之”字形路线。而蔗糖和磷酸根不易透过膜,走的是直线,因此显得运动速度比水快 ②在运输过程中,也可能有某种类似“层析”的机制使得各种溶质通过的速度不同 ③筛管分子是活细胞,一些溶质在运输过程中被代谢或相互影响从而改变运输速度。例如磷酸根在代谢上要比蔗糖活泼得多。 将茎表面擦伤,施用荧光染料,染料会被吸进韧皮部。经过一段时间后,在施用部位的上、下方取茎切片,发现染料向上和向下运输;在茎的一个节上的叶片上施用荧光染料,在另一个节的叶片上施用14CO2,用蚜虫吻针法在两个节之间收集溢泌液,发现其中含有两种示踪物质。这似乎明确地显示在一个筛管中存在双向运输。但这也可以作其他解释:首先,蚜虫口针法收集溢泌液通常需要数小时,在这样长的时间内,同一筛管内的运输方向可能发生了改变;其次,在筛管之间、甚至在韧皮部与木质部之间可以进行侧向转运。 以11CO2或14CO2作脉冲标记的实验表明,在单一筛管分子中,同化物运输是单向的。 早期的电镜观察表明,筛孔是被P-蛋白堵塞的,这一结果导致20世纪60年代对压力流假说的极大怀疑,并由此提出了一些新的假说。但随着实验技术的改进,研究人员证明以前的观察只是一种假象:当材料被切开或被缓慢固定时,在筛管内正的压力作用下,P-蛋白便堆积在筛孔上。如果将整株材料先在液氮中迅速冷冻,再进行化学固定,或在取样固定前先使植株萎蔫,以减小筛管中压力。观察到筛孔是开放的。 在一些实验中测定出的压力都足以使集流保持一定速度。例如柳树筛管中的压力可达1.0MPa;大豆源与库间的压力差为0.41MPa。而根据计算,维持集流所需要的压力差是0.12~0.46MPa。 可以用间接法测定韧皮部的压力:测定出水势和韧皮部汁液的渗透势,便可以计算出压力势。也可以直接测定筛管的压力:例如将一玻璃毛细管的一端封死,开口端与刺入植株筛管的蚜虫口针紧密连接,筛管汁液将被压入毛细管。通过测定毛细管中空气被压缩的程度,可以计算出筛管压力的大小。
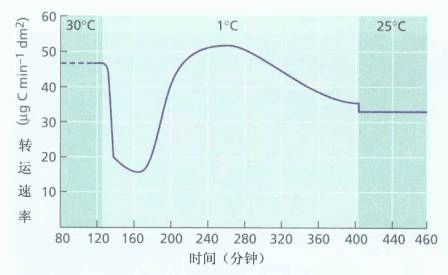
图6-16 甜菜韧皮部运输对低温的反应 源叶饲喂14CO2,20cm长的一段叶柄冷却至1℃,监测14CO2到达叶的速率以确定运输速率。实验中运输速率的暂时降低有可能是因冷却使运输的阻力增大(如增大了汁液的粘滞度等),而源叶通过装载机制建立起更高的压力后,运输速率可恢复。 在韧皮部运输的整个过程中,涉及能量消耗的部位有两处:一是源端光合同化物的韧皮部主动装载,以保持筛管内高的膨压;另一是库端韧皮部同化物的卸出与利用。因此,凡降低源库处韧皮部细胞代谢活性或能量供应的处理,如缺氧、低温、代谢抑制剂等,因降低源端同化物向筛管的装入和库端细胞对同化物的吸收利用而影响运输速率。然而,限制运输途径中的能量供应并不明显影响运输速度。例如给甜菜源叶的叶柄以1℃的处理,虽然呼吸被抑制90%(从而ATP的产生也同样被抑制),但运输速率在短暂受影响后又可恢复原水平(图6-16)。另外,茎或叶柄中的伴胞体积比筛细胞小,胞质稀;而源库端的伴胞或薄壁细胞体积比筛细胞大、胞质浓。从两者的结构比较分析中也可推测长距离运输的途中只需要少量能量,这些少量能量可能不是用于物质运输,而是用于维持韧皮部的生命活动。 上述的实验证据都支持压力流学说。从这里我们也看到了在科学发展过程中,一个学说是如何被提出,又是如何受到检验的,在科学研究中合理的设计、对实验结果做出正确的解释又是何等重要。 同化物韧皮部运输的研究已经历了70多年,尽管尚有许多方面需要深入研究,但目前普遍认为“压力流学说”是最能解释同化物韧皮部运输现象的一种理论。然而,必须指出的是,上述讨论的是被子植物中的情况。由于裸子植物韧皮部的结构与被子植物有很大的差异,因此,可以预测裸子植物同化物的韧皮部运输机理与被子植物的运输机理必然有许多不同之处,但这方面的研究尚很缺乏。 |