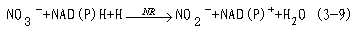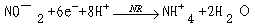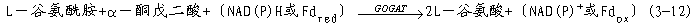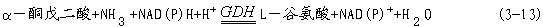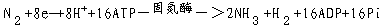|
受欢迎度:
第四节 氮的同化
一、植物的氮源
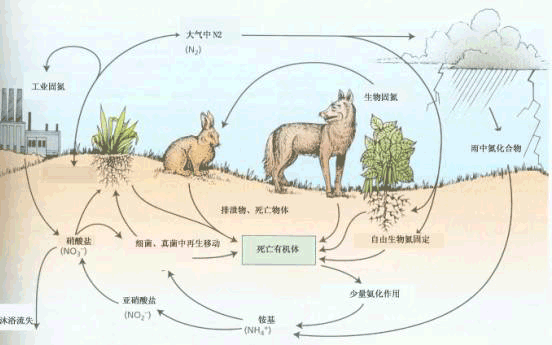
自然界中N素循环
空气中含有近79%的氮气(N2),然而,植物无法直接利用这些分子态氮。只有某些微生物(包括与高等植物共生的固氮微生物)才能利用大气中的氮气,而植物所利用的氮源,主要来自土壤。土壤中的有机含氮化合物主要来源于动物、植物和微生物躯体的腐烂分解,然而这些含氮化合物的大多是不溶性的,通常不能直接为植物所利用,植物只可以吸收其中的氨基酸、酰胺和尿素等水溶性的有机氮化物。尿素虽是植物良好的氮源,但由于它易被分解为NH3和CO2,所以施入田间的尿素,也只有一部分是以尿素分子的形式被植物吸收的。植物的氮源主要是无机氮化物,而无机氮化物中又以铵盐和硝酸盐为主,它们约占土壤含氮量的1-2%。植物从土壤中吸收铵盐后,可直接利用它去合成氨基酸。如果吸收硝酸盐,则必须经过代谢还原(metabolic reduction)才能被利用,因为蛋白质的氮呈高度还原态,而硝酸盐的氮则呈高度氧化态。 |
二、硝酸盐的还原
大多数植物虽能吸收NH 4,但在一般田间条件下,NO-3是植物吸收的主要形式。NO-3进入细胞后,就被硝酸还原酶和亚硝酸还原酶还原成铵。在此还原过程中,每形成一个分子NH 4要求供给8个电子。硝酸盐还原(nitrate reduction)的过程如下:
植物细胞硝酸盐同化,包括硝酸盐的跨质膜运输,然后经两步还原为氨
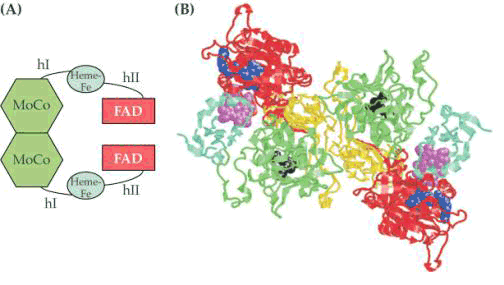
硝酸还原酶(nitrate reductase, NR)催化硝酸盐还原为亚硝酸盐。真核生物的NR由于来源不同,其亚基组成的数目和大小也不同,小的有2个亚基,大的可达8个亚基。现在认为硝酸还原酶是一种可溶性的钼黄素蛋白(molybdoflavoprotein),它由黄素腺嘌呤二核苷酸(FAD),细胞色素b557和钼复合体(Mo-Co)组成,分子量200 000~500 000。推测它的结构为同型二聚体(homodimer)(图3-18)。硝酸盐还原所需的供氢体是NADH(NADPH)。NO3-还原为NO2-的反应如下:
图3-18 高等植物硝酸还原酶的模型
A)硝酸盐还原酶的结构域结构。一个NR单体有三个主要的结构域,分别与钼辅因子、血红素和FAD相连。FAD连接区从NAD(P)H接受电子;血红素结构域运送电子到MoCo连接区,它传递电子给硝酸盐,hⅠ和hⅡ指铰链1和铰链2,分离功能结构域。(B)硝酸盐还原酶的条带图解。血红素辅基用紫色表示,FAD用蓝色表示,MoCo用黑色表示,2个单体之间的界面用黄色表示
硝酸还原酶是一种诱导酶(induced enzyme)。我国科学家吴相钰、汤佩松(1957)首先发现水稻幼苗若培养在含硝酸盐的溶液中就会诱导幼苗产生硝酸还原酶,如用不含硝酸盐的溶液培养,则无此酶出现。这也是国内外最早的有关高等植物体内存在诱导酶的报道。亚硝酸还原酶(nitrite reductase,NiR)催化亚硝酸盐还原为铵。在正常有氧条件下,由NR催化形成的亚硝酸盐,很少在植物体内积累,因为在植物组织中NiR的存在量大。亚硝酸还原酶分子量为61 000~70 000,它由两个亚基组成,其辅基由西罗血红素(sirohaem)和一个4Fe-4S簇组成。西罗血红素的作用是使6个电子转移到底物亚硝酸上,其反应如式3-10和图3-19所示:
图3-19亚硝酸还原的电子传递体系
Fdred和Fdox分别是还原态和氧化态的铁氧还蛋白
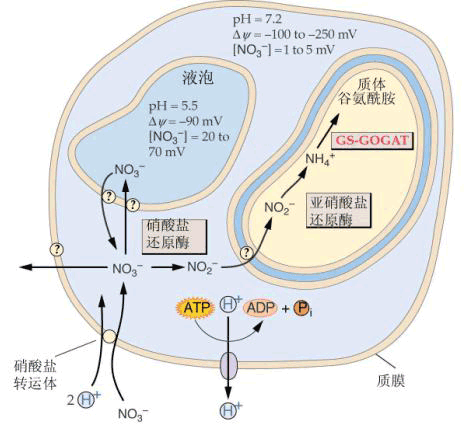
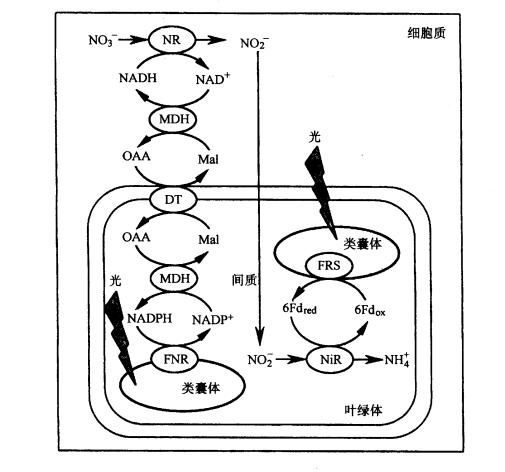
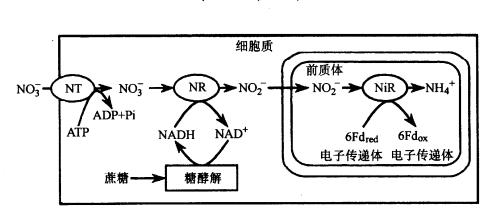
硝酸盐的还原可在根和叶内进行,通常绿色组织中硝酸盐的还原比非绿色组织中更为活跃。硝酸在叶内和根中的还原过程见图3-20和图3-21。
图3-20在叶中的硝酸还原
DT.双羧酸运转器; FNR.FdNADP还原酶; MDH:苹果酸脱氢酶;FRS.Fd还原系统
图3-21在根中的硝酸还原
NT.硝酸运转器
在绿叶中硝酸盐的还原是在细胞质中进行的,当硝酸盐被细胞吸收后,细胞质中的硝酸还原酶就利用NADH供氢体将硝酸还原为亚硝酸,而NADH是叶绿体中生成的苹果酸经双羧酸运转器,运送到细胞质,再由苹果酸脱氢酶催化生成的。NO-3还原形成NO-2后被运到叶绿体, 叶绿体内存在的NiR利用光合链提供的还原型Fd作电子供体将NO-2还原为NH 4。硝酸盐在根中的还原与叶中基本相同,即硝酸盐通过硝酸运转器进入细胞质,被NR还原为NO-2,但电子供体NADH来源于糖酵解。形成的NO-2再在前质体被NiR还原为NH 4。长期以来对根中存在的NiR的电子供体不清楚,但最近已从许多植物根中发现了类似Fd的非血红素铁蛋白或Fd-NADP还原酶。
硝酸盐在根部及叶内还原所占的比例受多种因素影响,包括硝酸盐供应水平、植物种类、植物年龄等。一般说来,外部供应硝酸盐水平低时,则根中硝酸盐的还原比例大。木本植物根的硝酸还原能力很强。作物中硝酸盐在根内还原能力依次为:燕麦>玉米>向日葵>大麦>油菜。此外根中硝酸盐的还原比例还随温度和植物年龄的增加而增大。通常白天硝酸还原速度显著较夜间为快,这是因为白天光合作用产生的还原力能促进硝酸盐的还原。
|
三、氨的同化
植物从土壤中吸收铵,或由硝酸盐还原形成铵后会立即被同化为氨基酸。氨的同化在根、根瘤和叶部进行,已确定在所有的植物组织中,氨同化是通过谷氨酸合成酶循环进行的(图3-22)。在这个循环中有两种重要的酶参与催化作用,它们分别是谷氨酰胺合酶(glutamine synthase,GS)和谷氨酸合成酶(glutamate synthetase,GOGAT)。
GS催化下列反应:
GS普遍存在于各种植物的所有组织中。它对氨有很高的亲和力,其Km为10-5 ~10-4mol·L-1,因此能防止氨累积而造成的毒害。谷氨酸合酶有两种形式,一是以NAD(P)H为电子供体的NAD(P)HGOGAT,另一是以还原态Fd为电子供体的FdGOGAT。两种形式的GOGAT均可催化如下反应:
NAD(P)H-GOGAT在微生物和植物中广泛存在,而Fd-GOGAT几乎存在于所有具有光合作用的生物体中。被子植物的组织都有较高的GOGAT活力,且种子、根及叶片等组织中两种GOGAT共存。一般叶片中的酶活力比根中的高,其中FdGOGAT活力又较NAD(P)HGOGAT活力高(种子中例外)。此外,还有谷氨酸脱氢酶(glutamate dehydrogenase, GDH)也参与氨的同化过程,它催化下列反应:
谷氨酸脱氢酶在植物同化氨的过程中不很重要,因为GDH与NH3的亲和力很低,其Km值为5.2~7.0mmol·L-1 。但GDH在谷氨酸的降解中起了较大作用,GDH主要存在于根和叶内的线粒体中,而叶绿体中的量很少。三种酶在细胞中的定位,经研究已被确认:绿色组织中GOGAT存在于叶绿体内;GS在叶绿体和细胞质中都有存在,而GDH则主要存在于线粒体中。在非绿色组织,特别是根中,GS和GOGAT定位于质体,GDH定位在线粒体中,而GS是否存在于细胞质中还有争论。 界中的氮素平衡,具有十分重大的意义。
图3-22 谷氨酸合成酶循环
|
四、生物固氮
在一定条件下,N2可与其它物质进行化学反应,固定形成氮化物,这个过程称为固氮作用。在自然固氮中,约有10%是通过闪电完成的,其余90%是通过微生物完成的。某些微生物把空气中的游离氮固定转化为含氮化合物的过程,称为生物固氮(biological nitrogen fixation)。生物固氮对农业生产和自然界中的氮素平衡,具有十分重大的意义。
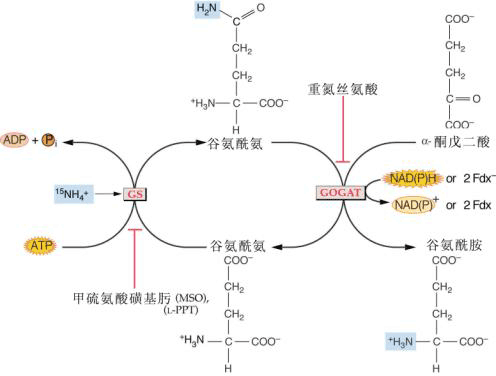
大豆根瘤菌,许多节结瘤是Rhizobium japinicum感染的结果
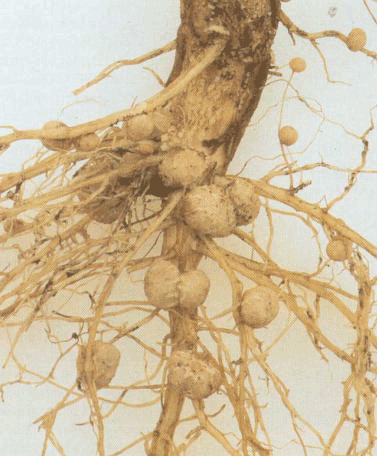
图 3-23固氮酶催化反应
铁氧还蛋白还原铁蛋白,与ATP结合,铁蛋白还原钼铁蛋白,最后还原N2成为NH3
生物固氮是由两类微生物来实现的。一类是自生固氮微生物包括细菌和蓝绿藻,另一类是与其它植物(宿主)共生的微生物,例如与豆科植物共生的根瘤菌,与非豆科植物共生的放线菌,以及与水生蕨类红萍(亦称满江红)共生的蓝藻(鱼腥藻)等,其中以根瘤菌最重要。固氮微生物体内含有固氮酶(nitrogenase),它具有还原分子氮为氨的功能。一般认为固氮酶是由钼铁蛋白和铁蛋白构成的复合物。两者都是可溶性蛋白质。氨是生物固氮的最终产物,分子氮被固定为氨的总反应式如下:
在整个固氮过程中,以铁氧还蛋白为电子供体,去还原铁蛋白,后者进一步与Mg·ATP结合,形成还原型的Mg·ATP铁蛋白,电子接着流到钼铁蛋白,还原型的钼铁蛋白接着还原N2,最终形成NH3(图3-23)。固氮酶也可以还原质子(H )而放出氢(H2)。氢在氢化酶(hydrogenase)作用下,又可还原铁氧还蛋白,这样氮的还原就形成一个电子传递的循环。生物固氮虽然可以利用固氮酶作用得到可利用的氮来满足作物需要。但是从3-14式可见,固氮酶固定1分子N2s要消耗8个e-和16个ATP。据计算,高等植物固定1g N2要消耗有机碳12g。看来,如何减少固氮所需的能量投入量是生物固氮研究中凾待解决的问题。
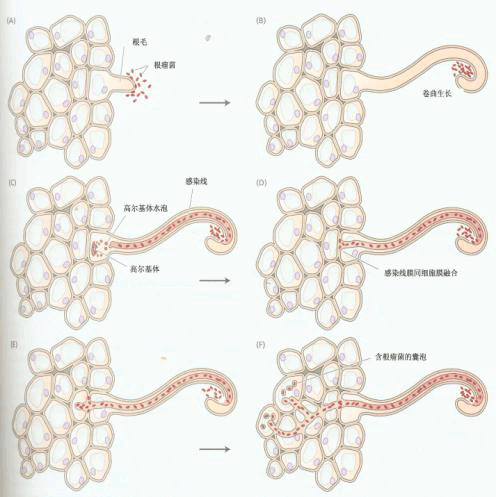
根瘤有机合成中根瘤菌感染过程
(A)随植物注射化学诱导剂后,根瘤菌结合刚形成的根毛。(B)随细菌产生影响因子,根毛呈现弯曲生长,根瘤菌在根毛圈内增生扩散。(C)根毛壁的局部降解导致根细胞中高尔基体小泡感染形成感染线。(D)感染线达到细胞终点后,它的膜同根毛细胞膜融合生长。(E)根瘤菌在质外体中释放,渗透穿过胞间层达到质膜的亚表皮细胞,导致激发端开口与第一条感染线相通的新感染线的形成。(F)感染线不断伸长分枝直到目标细胞,在那里囊泡构成的植物膜将释放到细胞质中的细菌细胞包围。
|
|